Welcome to our R Shiny Application!
In the Mediterranean climate of the Pacific Northwest, dry summers result in prolonged periods of low flows in streams. Climate change, habitat alterations, and increasing water demands are leaving less water available for streams. As water levels drop, some small streams become fragmented, transforming from a ribbon of continuous habitat into a series of isolated pools. Fragmented streams pose a serious threat to salmon; juveniles that become stranded in small pools are at increased risk to overheat, starve, or be consumed by predators.


Healthy salmon populations can cope with fragmentation and recover from a bad drought-year. However, for depressed populations (e.g., ESA-listed), there is a strong need for adaptation strategies that reduce the risks of drought-induced fragmentation. While a loud and high profile debate has ensued over the permanent translocation of species (i.e., assisted migration), there has been much less attention given to the use of temporary translocation to buffer species from seasonal risks. One form of temporary translocation is known as fish rescue, which seeks to reduce drought related mortality in wild fish by manually moving individuals from fragmented areas to artificial rearing facilities that provide refuge during periods of low flow. Fish rescue differs from the more typical fish salvage operations where individuals are instead moved to free-flowing habitat.
This Shiny application explores how seasonal fish rescue affects the population dynamics of coho salmon. This application considers fish rescue that involves seasonal or annual translocation to captive rearing. Before using this application, we strongly recommend that you first read Brittany Beebe's 2019 Master's Thesis from Oregon State University entitled Evaluating Fish Rescue as a Drought Adaptation Strategy for Imperiled Coho Salmon: A Life-Cycle Modeling Approach (Link). This document provides a full description of the life-cycle model and additional background on fish rescue and ecological drought. The thesis document also provides discussion of the potential costs and benefits associated with fish rescue.
Understanding Model Mechanics
The model framework reflects the basic life cycle of coho salmon and simulates abundance across five serial life stages: spawner, fry, parr, smolt, and adult. Wild fish move sequentially through each life stage and are subjected to corresponding survival processes. Fish rescue is simulated as an alternative pathway between the fry and smolt life stages. The model explores two captive rearing durations: long- and short-term rescue. For long-term rescue, fish are held for roughly a year through the fry-to-smolt stages. This transition encompasses both summer and winter survival. For the short-term rescue, fish are held in captivity only during the season of wetted habitat contraction, which was considered as the summer fry-to-parr survival stage.
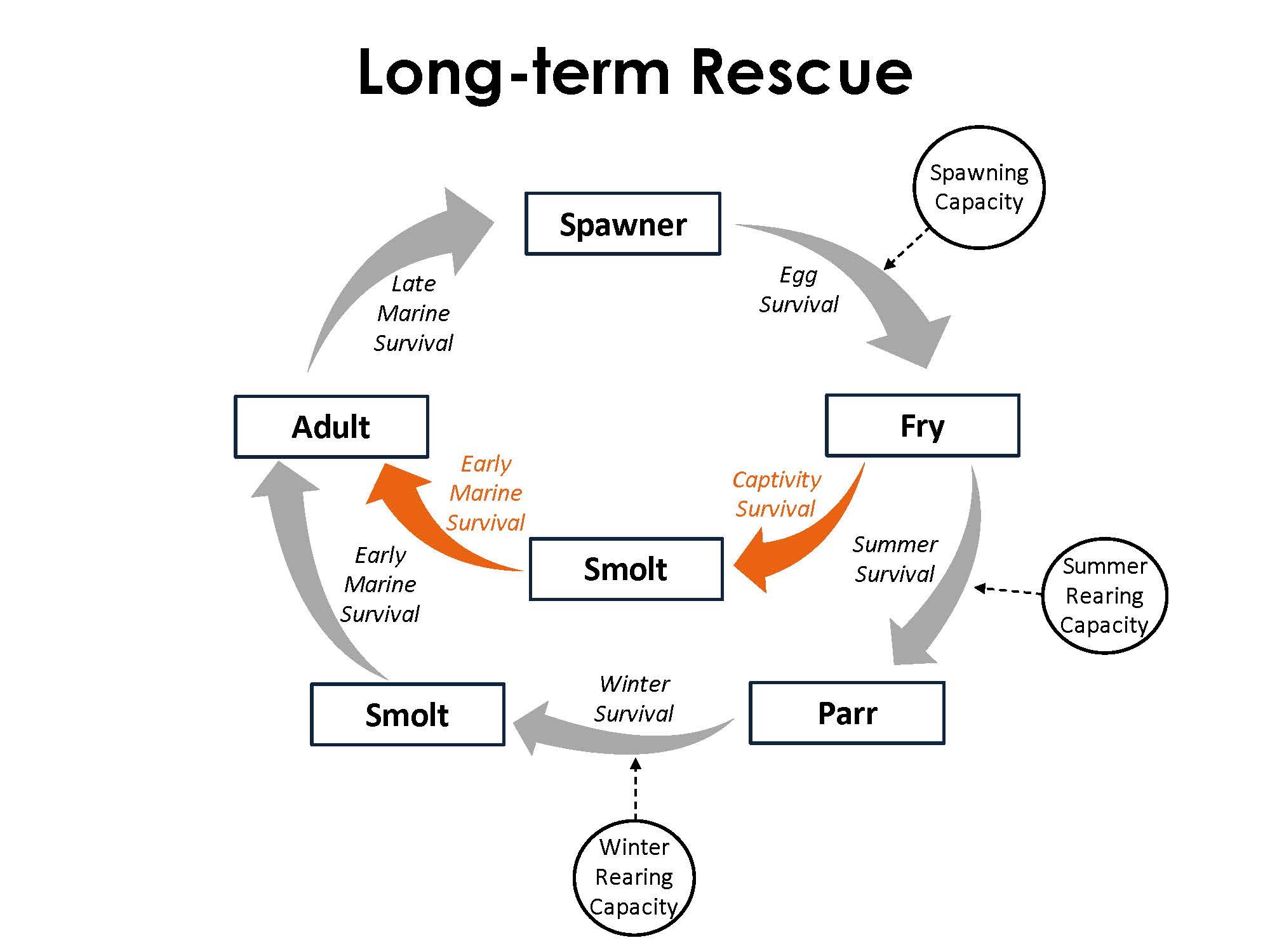
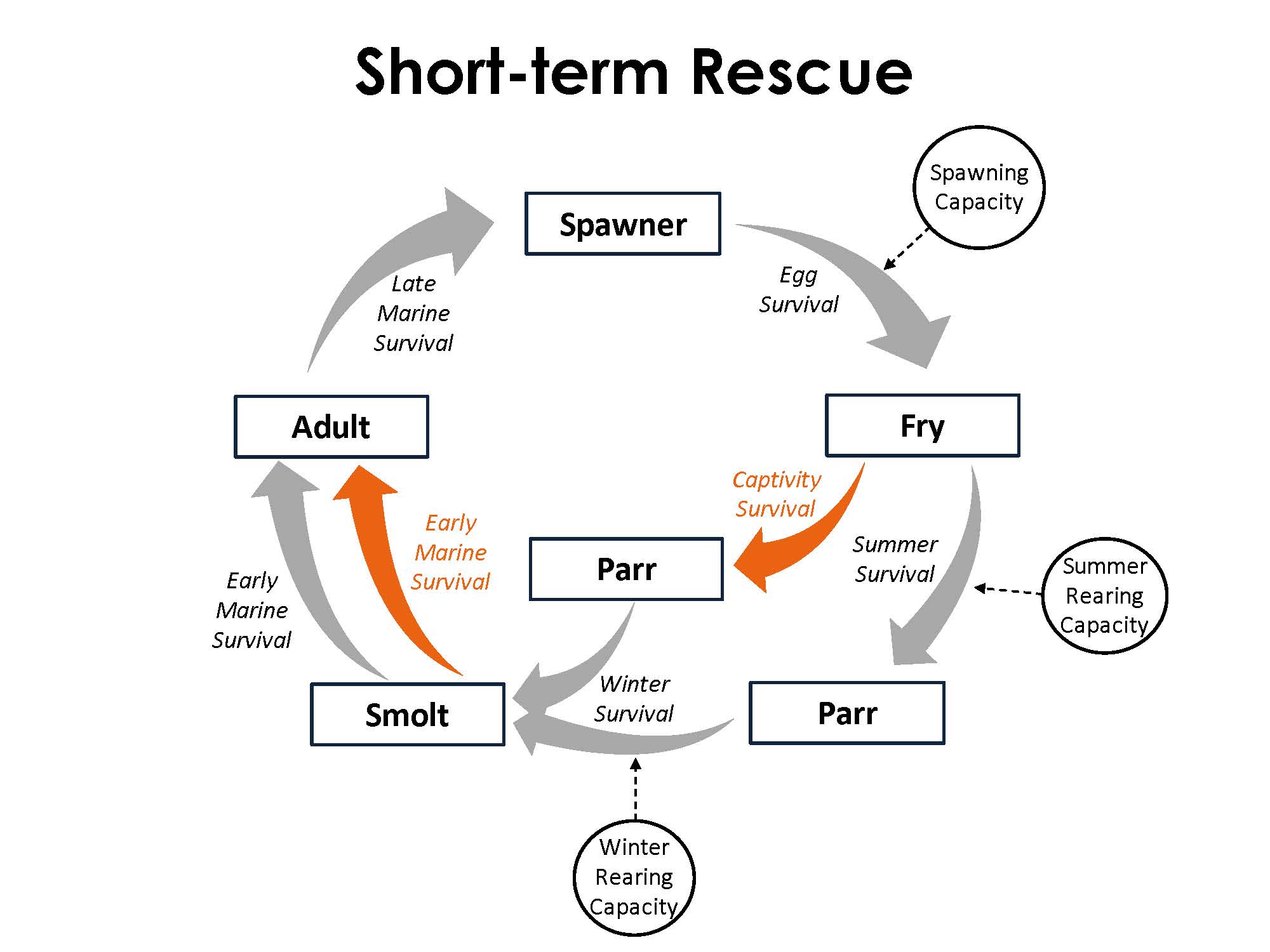
Survival
Survival is represented by the transition of individuals from one life stage to the next. The transition between the spawner and fry life stages includes fecundity in addition to survival. Three general methods were used to calculate survival. The Beverton-Holt function is used for freshwater survival (Moussalli & Hilborn 1986), a beta distribution is used for marine survival (Hill et al. 2003), and a constant parameter value is used for survival in captivity (NWFR, personal communication). The survival estimates calculated for each transition are used in a binomial distribution to incorporate demographic stochasticity to the simulated number of individuals at the subsequent life stage (Nickelson & Lawson 1998).
Rescue
Although not bred in captivity, rescued fish spend much of their freshwater life stages in artificial conditions, and we assumed this could negatively affect smolt-to-adult return rates. Reported estimates of marine survival of hatchery fish compared to wild fish range from ~37-100% (Jonsson et al. 2003; Kallio-Nyberg et al. 2004; Jokikokko et al. 2006; Hyvarinen & Rodewald 2013). In the model, early marine survival of rescued fish is calculated by multiplying early marine survival of wild fish by a marine survival penalty value.
The attempted rescue level remains constant for every generation of a given simulation. Since fry abundance may fall below the attempted rescue level, a capped linear functional response (i.e., hockey-stick) is used in which a percentage of the total fry are captured up to the attempted rescue level. In other words, when fry abundance is less than the captive rearing capacity, an encounter probability is used. This avoids rescuing an unrealistic percentage of available fry.
Drought
Drought conditions are explored by adjusting the summer rearing capacity. To explore more severe drought, set the summer capacity to a lower value.
Mean Smolt Abundance Over Time
Mean Spawner Abundance Over Time
Extinction Risk Over Time
Mean Spawner Abundance Across Drought Conditions
Extinction Risk Across Drought Conditions
Model Settings
- Number of simulations: The number of simulations to run. Default = 3,000 simulations
- Simulation length: The length of time each simulation is run. Default = 33 generations (i.e., 99 years)
- Initial spawner abundance: The initial abundance of total spawners (male plus female). Default = 300 spawners
- Extinction threshold: A value of spawner abundance (male plus female) below which extinction is likely. Default = 50 spawners
Rescue Parameters
- Holding time: The length of time that rescued fish are held in captivity. Long-term implies rescued fish are held over both summer and winter. Short-term implies rescued fish are held only over summer. Default = Long-term
- Number of fry rescued: The number of fry rescued for each rescue period. Default = 10,000
- Encounter probability: The probability of encountering fry when abundance is lower than number of fry to be rescued. This avoids rescuing all fish when fry abundance is low. Default = 0.4
- Captivity survival rate: Rescued fish survival spanning from the time of stream removal in early summer until the time of release in fall for short-term rescue or the following spring for long-term rescue. Default = 0.98 (Source: personal communication with Northwest Wild Fish Rescue)
- Marine survival penalty: Penalty applied to early marine survival for rescued fish. The value selected represents survival of rescued fish compared to survival of wild fish. A lower value indicates lower survival and thus a greater penalty for being rescued. For example, a value of 40% indicates that rescued fish survival is 40% of the wild fish survival (i.e., rescued fish survival = 40% of wild fish survival). Default = 60%
Freshwater Rearing Capacities
- Spawning capacity: The maximum number of eggs sustained by the stream. Calculated from multiplying stream length (km) by maximum spawner density (19 females/km). Default = 902,500 (Source: Bradford et al. 2000)
- Summer capacity: The maximum number of fry sustained by the stream when in its most contracted state during the summer. This parameter is used as a proxy for drought condition. A low number indicates more severe drought, while a larger number indicates less severe drought. Default = 10,000
- Winter capacity: The maximum number of fry sustained by the stream during highest winter flows. Default = 20,000
Reproduction
- Fecundity: The number of eggs produced by each female spawner. Default = 2500 eggs/female
- Spawner sex ratio: The percent of total spawners that are female. Default = 50% female (i.e., 1:1)
Density Independent Survival (Max Productivity)
- Egg to fry survival: Survival from egg to fry as density approaches zero. Default = 0.429 (Source: Nickelson 1998)
- Oversummer survival: Oversummer survival of wild fish from fry to parr as density approaches zero. Default = 0.26 (Source: Nickelson 1998)
- Overwinter survival: Overwinter survival of wild fish from parr to smolt as density approaches zero. Default = 0.9 (Source: Nickelson 1998)
Early Marine Survival for Wild Fish
- Mean: Mean marine survival for wild fish from outmigration through first year in marine environment. Default = 0.098 (Source: Bradford 1995)
- Standard deviation: Standard deviation for early marine survival for wild fish. Default = 0.01
Late Marine Survival for All Fish
- Mean: Mean late marine survival for all fish (wild and rescued) Default = 0.15
- Standard deviation: Standard deviation for late marine survival for all fish. Default = 0.01
Barrowman NJ, RA Myers, R Hilborn, DG Kehler, and CA Field. 2003. The variability among populatinos of coho salmon in the maximum reproductve rate and depensation. Ecological Applications 13:784-793.
Beebe. 2019. Evaluating Fish Rescue as a Drought Adaptation Strategy for Imperiled Coho Salmon: A Life-Cycle Modeling Approach. Master's Thesis. Oregon State University, Corvallis, Oregon. Link
Bradford. 1995. Comparative review of Pacific salmon survival rates. Canadian Journal of Fisheries and Aquatic Sciences 52:1327-1338.
Bradford MJ, RA Myers, and JR Irvine. 2000. Reference points for coho salmon (Oncorhynchus kisutch) harvest rates and escapement goals based on freshwater production. Canadian Journal of Fisheries and Aquatic Sciences 57:677-686.
Hill MF, LW Botsford, and A Hastings. 2003. The effects of spawning age distribution on salmon persistence in fluctuating environments. Journal of Animal Ecology 72:736-744.
Hyvarinen P and P Rodewald. 2013. Enriched rearing improves survival of hatchery-reared Atlantic salmon smolts during migration in the River Tornionjoki. Canadian Journal of Fisheries and Aquatic Sciences 70:1386-1395.
Jokikokko E, I Kallio-Nyberg, I Saloniemi, and E Jutila. 2006. The survival of semi-wild, wild and hatchery-reared Atlantic salmon smolts of the Simojoki River in the Baltic Sea. Journal of Fish Biology 68:430-442.
Jonsson N, B Jonsson, and LP Hansen. 2003. The marine survival and growth of wild and hatchery-reared Atlantic salmon: growth and survival of salmon. Journal of Applied Ecology 40:900-911.
Kallio-Nyberg I, E Jutila, I Saloniemi, and E Jokikokko. 2004. Association between environmental factors, smolt size and the survival of wild and reared Atlantic salmon from the Simojoki River in the Baltic Sea. Journal of Fish Biology 65:122-134.
Moussalli E & R Hilborn. 1986. Optimal Stock Size and Harvest Rate in Multistage Life History Models. Canadian Journal of Fisheries and Aquatic Sciences 43:135-141.
Nickelson TE. 1998. A habitat-based assessment of coho salmon production potential and spawner escapement needs for Oregon coastal streams. Oregon Department of Fish Wildlife Fish Division Information Report 98-4.
Nickelson TE and PW Lawson. 1998. Population viability of coho salmon, Oncorhynchus kisutch, in Oregon coastal basins: application of a habitat-based life cycle model. Canadian Journal of Fisheries and Aquatic Sciences 55:2383-2392.
Steel EA, DW Jensen, KM Burnett, K Christiansen, JC Firman, BE Feist, KJ Alauf, and DP Larson. 2012. Landscape characteristics and coho salmon (Oncorhynchus kisutch) distributions: explaining abundance versus occupancy. Canadian Journal of Fisheries and Aquatic Sciences 69:457-468.